


Dans cette fiche explicative, nous allons apprendre à décrire la structure et la fonction des membranes, et comment les facteurs affectant la perméabilité des membranes peuvent être étudiés.
Tout organisme vivant, qu’il soit eucaryote/procaryote, animal ou bactérie, possède des cellules entourées par une membrane. Ces membranes servent de gardiennes pour la cellule, en contrôlant ce qui entre et sort, entre autres rôles importants.
Les cellules eucaryotes ont même des membranes à l’intérieur des cellules elles-mêmes ! Ces membranes intracellulaires délimitent certains organites eucaryotes ; elles permettent à différentes conditions de coexister, donc différentes réactions chimiques peuvent se produire dans chaque zone distincte de la cellule. Cette séparation des composants d’une cellule est appelée compartimentation. La compartimentation des organites dans une cellule peut être comparée à la manière dont les pièces d’une maison sont séparées les unes des autres.
Définition : Compartimentation
La compartimentation est la formation de différentes zones délimitées par une membrane dans une cellule, pour permettre à des réactions incompatibles d’avoir lieu dans différentes parties de la cellule.
Un organite est une structure subcellulaire qui remplit une fonction spécifique. Il peut être délimité ou non par une membrane.
Définition : Organite
Un organite est une structure subcellulaire qui remplit une fonction spécifique.
La membrane qui délimite un organite peut avoir une ou deux couches, auquel cas on parle de double membrane. Parmi les organites à double membrane, on retrouve notamment le noyau, les mitochondries et les chloroplastes. Parmi ceux à une seule membrane, il y a les lysosomes et l’appareil de Golgi.
Les membranes intracellulaires des organites eucaryotes sont aussi souvent le site de réactions chimiques. Le réticulum endoplasmique rugueux (RER) en est un exemple, les ribosomes incrustés dans sa membrane permettant la synthèse des protéines. C’est pour ça que le RER est « rugueux » comparé au réticulum endoplasmique lisse (REL), qui possède également une double membrane, mais pas de ribosomes. Le RER et le REL ont tous deux une seule membrane. La membrane interne des mitochondries et les membranes des chloroplastes sont les sites de la respiration aérobie et de certaines réactions de photosynthèse respectivement.
Exemple 1: Identifier les organites délimités par des membranes et décrire leur fonction
Les cellules eucaryotes contiennent de nombreux organites possédant des membranes internes.
- Quel organite parmi les suivants n’a pas de membrane dans sa structure ?
- le ribosome
- l’appareil de Golgi
- le réticulum endoplasmique
- le noyau
- le lysosome
- Les membranes internes créent des zones à conditions différentes pour que des réactions spécifiques aient lieu dans la cellule.
Quel terme décrit cette adaptation structurelle des cellules eucaryotes ?
Réponse
Partie 1
En plus des membranes entourant la cellule, de nombreux organites des cellules eucaryotes ont également des membranes. Un organite est une structure subcellulaire qui remplit une fonction spécifique.
Les organites peuvent être entourés d’une ou deux membranes, ou ne pas en avoir du tout. Par exemple, les ribosomes et les centrosomes n’ont pas de membrane plasmique, tandis que les lysosomes, le réticulum endoplasmique et l’appareil de Golgi ont une membrane simple et que le noyau et les mitochondries ont une double membrane !
Par conséquent, l’organite de la liste qui n’a pas de membrane est le ribosome.
Partie 2
Les membranes qui entourent certains organites séparent leur contenu du milieu environnant, permettant à différentes conditions d’être présentes à l’intérieur de chaque organite. Cela signifie que différentes réactions chimiques peuvent avoir lieu dans les différents organites de la cellule, mais aussi dans les membranes elles-mêmes.
Un exemple en est le réticulum endoplasmique rugueux (RER). Le RER possède à sa surface des ribosomes, qui effectuent la synthèse des protéines, mais n’ont pas de membranes. La séparation du contenu d’un organite de son environnement extérieur par une membrane est appelée la compartimentation.
Par conséquent, le terme qui décrit l’adaptation structurelle des membranes internes dans les cellules eucaryotes est la compartimentation.
Suivant leur emplacement et leur composition, les membranes ont tout un éventail de fonctions, mais leur point commun à toutes est qu’elles permettent de séparer une cellule, ou différentes zones d’une cellule, de leur environnement.
Les membranes plasmiques sont souvent décrites comme semi-perméables, ce qui signifie que seules certaines molécules peuvent les traverser grâce à des protéines à leur surface. Les membranes peuvent être comparées aux murs séparant les pièces d’une maison. Elles contrôlent les substances qui entrent et sortent de la cellule, grâce à des protéines agissant comme les « portes » de chaque « chambre », qui peuvent être « ouvertes » ou « fermées » pour des substances spécifiques.
Qu’elles soient internes ou situées à la surface de la cellule, toutes les membranes plasmiques ont la même structure de base que nous allons étudier avant d’apprendre leurs fonctions et les facteurs qui affectent leur perméabilité.
Définition : Membrane plasmique
Une membrane plasmique ou cellulaire est une barrière semi-perméable constituée d’une bicouche de phospholipides et d’autres molécules intégrées. Les membranes plasmiques entourent toutes les cellules, les séparent de leur environnement externe et compartimentent certains organites au sein des cellules eucaryotes.
Toutes les membranes, comme par exemple la membrane plasmique d’une cellule animale classique en figure 1, sont constituées d’une structure appelée la bicouche lipidique.

Décomposons le terme bicouche lipidique pour mieux le comprendre : les membranes sont principalement composées de molécules appelées phospholipides, qui ont une tête polaire hydrophile (attirée par l’eau) et une queue d’acide gras hydrophobe (repoussée par l’eau).
Le mot bicouche se réfère au fait que ces phospholipides forment deux couches, l’une orientée vers l’environnement extérieur, et l’autre vers le contenu du cytoplasme (voir figure 1). Les têtes polaires hydrophiles sont attirées par les molécules d’eau.
Comme la plupart des cellules et leurs composants sont en milieu aqueux (contenant de l’eau), les têtes polaires hydrophiles s’organisent de sorte à être proches de l’eau aux surfaces interne et externe de la membrane. Les queues d’acides gras sont cependant repoussées par les molécules d’eau (hydrophobes) et se tournent vers l’intérieur et l’une vers l’autre, comme le montre la figure 1.
Il est utile de noter que les queues d’acides gras hydrophobes dans la bicouche lipidique peuvent être constituées d’acides gras saturés ou insaturés. Vous pouvez voir un acide gras saturé et un acide gras insaturé dans la queue de la molécule de phospholipide sur la figure 2 ci-dessous.

Vous avez peut-être remarqué que les structures des acides gras saturé et insaturé qui composent la queue de ce phospholipide sont différentes. Les acides gras saturés sont composés de molécules de carbone reliées par des liaisons simples, formant une chaîne linéaire de carbones complètement saturés par des atomes d’hydrogène. Les acides gras insaturés ont au moins une double liaison entre deux atomes de carbone, ce qui forme un « coude » dans la queue, et ne sont donc pas complètement saturés en atomes d’hydrogène.
Ces coudes signifient que les queues d’acides gras adjacentes ne peuvent pas être aussi serrées que les queues d’acides gras saturés, ce qui augmente la fluidité de la membrane plasmique.
Définition : Bicouche lipidique
La bicouche lipidique est une membrane constituée de deux couches de phospholipides, avec une tête polaire hydrophile et des queues d’acides gras hydrophobes.
Terme clé : Tête polaire (hydrophile)
Les têtes polaires (aussi dites hydrophiles) sont les parties des phospholipides qui sont attirées par l’eau, et constituent donc la surface des membranes plasmiques dans les milieux aqueux.
Terme clé : Queue d’acide gras hydrophobe
Les queues d’acides gras hydrophobes sont les parties des phospholipides qui sont repoussées par l’eau, et forment donc le cœur interne des membranes plasmiques.
Exemple 2: Décrire la structure des membranes biologiques
Quel énoncé parmi les suivants décrit correctement la structure des membranes biologiques ?
- Les membranes cellulaires sont formées de deux couches de phospholipides, où les têtes hydrophiles sont tournées vers l’extérieur.
- Les membranes cellulaires sont formées d’une seule couche de phospholipides, reliés par leurs têtes hydrophobes.
- Les membranes cellulaires sont formées de deux couches de phospholipides, où les queues hydrophiles se lient dans une matrice.
- Les membranes cellulaires sont formées d’une couche de phospholipides et d’une couche de glycolipides, reliées par des queues hydrophobes.
Réponse
Les membranes cellulaires sont décrites comme des bicouches de phospholipides. Cela signifie qu’elles se composent de deux couches de phospholipides adjacentes. Les cellules se trouvent généralement dans un milieu aqueux, contenant des molécules d’eau, et le cytoplasme est également aqueux. Cela signifie que les têtes hydrophiles des phospholipides sont tournées vers l’extérieur pour s’associer aux molécules d’eau dans les environnements extérieur et intérieur de la cellule. Les queues d’acides gras des phospholipides sont hydrophobes, donc elles sont repoussées par les molécules d’eau dans le cytoplasme et à l’extérieur de la cellule, et sont tournées vers l’intérieur.

Par conséquent, l’énoncé décrivant correctement la structure de la membrane de surface cellulaire est le A : les membranes cellulaires sont formées de deux couches de phospholipides, où les têtes hydrophiles sont tournées vers l’extérieur.
Lorsqu’elle a été observée pour la première fois au microscope électronique, la bicouche lipidique ressemblait à des « rails de train », ce qui a aidé les scientifiques à déterminer que la membrane possédait deux couches. Vous comprendrez mieux pourquoi on parlait de « rails de train » en regardant la micrographie en fausses couleurs représentant les bicouches lipidiques entre quatre cellules épithéliales ci-dessous.
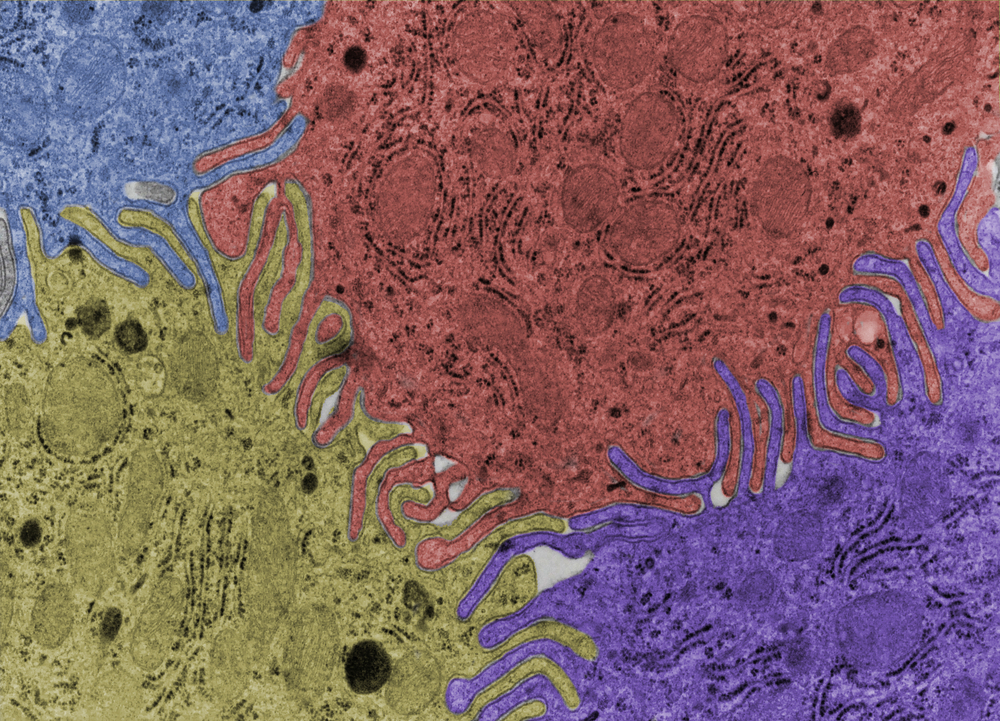
Cette observation a aidé les scientifiques à développer le modèle de la mosaïque fluide, décrivant comment les phospholipides sont, dans une certaine mesure, libres de se déplacer de manière fluide dans la membrane, car ils ne sont pas chimiquement liés les uns aux autres, mais simplement attirés ou repoussés par les molécules d’eau.
Le terme mosaïque se réfère aux molécules (principalement des protéines) de différentes formes et tailles qui sont intégrées dans diverses sections de la membrane plasmique, un peu comme les carreaux d’une mosaïque. Nous allons maintenant discuter du rôle de ces différentes molécules « mosaïques », visibles sur la figure 4 ci-dessous.
Terme clé : Modèle de la mosaïque fluide
Le modèle de la mosaïque fluide permet de comprendre le fonctionnement des membranes plasmiques : les phospholipides sont fluides et se déplacent librement dans la membrane, et des molécules de différentes formes et tailles sont intégrées dans la membrane comme des carreaux de mosaïque.

Diverses protéines sont intégrées dans la bicouche lipidique. Il y a notamment des protéines dites intrinsèques (ou transmembranaires ou intégrales), qui traversent les deux couches de la membrane, comme sur la figure 4. Toutes les protéines intrinsèques sont des protéines globulaires possédant des régions hydrophobes qui s’intègrent à la membrane.
L’exemple de la figure 4 est un canal ionique. Ces protéines transmembranaires contiennent un pore hydrophile, que les molécules polaires et les ions traversent passivement par diffusion selon le gradient de concentration. Ces substances ne pourraient pas traverser la membrane autrement en raison de la nature hydrophobe des queues d’acides gras des phospholipides.
Les transporteurs membranaires sont un autre type de protéine intrinsèque, modélisé sur la figure 5. Ils contiennent des récepteurs spécifiques à une molécule.
Par exemple, sur la figure 5, le glucose est transporté de l’espace extracellulaire à l’intérieur de la cellule. En raison de la forme spécifique du récepteur, aucune autre molécule ne rentre dans le transporteur membranaire de glucose. Visualisez le glucose comme la seule molécule possédant une « clé » pour cette protéine « porte » spécifique.
Lorsque le glucose se lie au récepteur du transporteur membranaire, la protéine change de forme et libère le glucose de l’autre côté de la membrane. Suivant la concentration en glucose à l’intérieur et à l’extérieur de la cellule, les transporteurs membranaires peuvent effectuer un transport actif ou passif.
Le transport actif nécessite de l’énergie pour faire traverser des molécules, car elles se déplacent contre leur gradient de concentration. Le transport passif ne nécessite pas d’énergie pour faire traverser les molécules, puisqu’elles se déplacent selon le gradient de concentration. Si la concentration en glucose à l’extérieur de la cellule est plus élevée qu’à l’intérieur, par exemple, le glucose entrera dans la cellule via les transporteurs membranaires par transport passif, selon le gradient de concentration, sans nécessiter d’énergie.

Terme clé : Protéines intrinsèques
Les protéines intrinsèques sont intégrées dans les deux couches d’une membrane et peuvent être soit des transporteurs membranaires de substances de manière active ou passive dans ou hors de la cellule, soit des canaux ioniques pour le transport passif uniquement.
Terme clé : Canaux ioniques
Les canaux ioniques sont des protéines intrinsèques qui créent un canal hydrophile permettant la diffusion de molécules et d’ions polaires à travers les membranes selon leur gradient de concentration.
Terme clé : Transporteurs membranaires
Les transporteurs membranaires sont des protéines intrinsèques possédant une forme spécifique complémentaire à certaines molécules, et lors de leur liaison, elles subissent un changement de forme pour transporter les molécules à travers une membrane contre (transport actif) ou selon (transport passif) leur gradient de concentration.
Les glycoprotéines sont un exemple de protéine intrinsèque, également visible sur la figure 4. Le préfixe glyco indique que la protéine est attachée à une chaîne glucidique (sucre). Les glycoprotéines ont deux utilités : elles aident les cellules à adhérer les unes aux autres et à se lier, et elles peuvent aussi servir de récepteurs.
Le rôle général d’un récepteur est de permettre à un élément chimique spécifique, tel qu’une hormone, de s’y lier et d’entraîner une réponse dans la cellule. C’est ce qu’on appelle la signalisation cellulaire, qui permet aux cellules du corps de communiquer efficacement.
Terme clé : Glycoprotéines
Les glycoprotéines sont des protéines intrinsèques attachées à une chaîne glucidique, qui permettent aux cellules d’adhérer les unes aux autres et de servir de récepteurs à différents éléments chimiques pour entraîner une réponse cellulaire (signalisation cellulaire).
Les glycolipides, également visibles sur la figure 4, sont semblables aux glycoprotéines mais, au lieu d’avoir une chaîne glucidique attachée à une protéine intrinsèque, ils s’attachent directement aux phospholipides dans la bicouche. On les appelle parfois marqueurs cellulaires ou antigènes, car ils sont utilisés par le système immunitaire dans la reconnaissance cellulaire pour identifier les cellules comme appartenant au soi ou non. Si les cellules n’appartiennent pas à l’organisme lui-même, elles sont étrangères et donc potentiellement nocives et doivent être détruites.
Définition : Glycolipides
Les glycolipides sont des lipides attachés à des chaînes glucidiques, souvent appelés marqueurs cellulaires ou antigènes, qui peuvent être reconnus par le système immunitaire comme appartenant au soi ou étrangers, ce qui indique s’ils sont attachés aux cellules d’un autre organisme tel qu’un agent pathogène.
Un exemple de glycolipides dans les cellules humaines sont les groupes sanguins ABO. Des glycolipides présents sur la membrane de surface des globules rouges agissent comme des antigènes, spécifiques à l’un des quatre groupes sanguins : A, B, AB et O.
Les protéines extrinsèques ou périphériques ne traversent pas toute la membrane, contrairement aux protéines intrinsèques, comme le montre la figure 4. À la place, elles sont intégrées d’un seul côté de la bicouche lipidique et interagissent avec la tête polaire d’un phospholipide à la surface interne ou externe de la membrane ou avec une protéine intrinsèque.
Définition : Protéines extrinsèques
Les protéines extrinsèques sont des protéines périphériques intégrées uniquement d’un côté de la membrane, qui interagissent avec la tête polaire d’un phospholipide ou avec des protéines intrinsèques.
La dernière structure dont nous allons discuter est le cholestérol, visible sur la figure 4 sous la forme d’une petite structure rouge intégrée entre les molécules de phospholipides. Le cholestérol est un lipide abondant dans les membranes des cellules animales. Il a une tête hydrophile et une queue hydrophobe rigide et volumineuse, qui interagit avec les queues des phospholipides dans la membrane. Grâce à ces interactions qui modulent l’organisation des phospholipides dans la membrane, le cholestérol peut augmenter ou réduire la fluidité des membranes en fonction de la température, comme nous allons le voir.
Terme clé : Cholestérol
Le cholestérol est une molécule lipidique qui régule la fluidité des membranes sur un large éventail de températures en s’insérant entre les molécules de phospholipides.
Exemple 3: Décrire le mouvement de molécules à travers la membrane plasmique
Le cyanure d’hydrogène est une petite molécule apolaire toxique libérée par certaines plantes pour repousser les herbivores. Le cyanure traverse les membranes et inhibe un processus clé de la respiration.

Par quel moyen le plus probable cette molécule traverse-t-elle les membranes ?
- Elle diffuse à travers la bicouche lipidique.
- Elle diffuse en utilisant un transporteur membranaire.
- Elle entre par un canal ionique.
- Elle entre par endocytose.
Réponse
Pour déduire la méthode par laquelle une molécule traverse une membrane plasmique, étudiez les données sur la molécule dans la question. On peut voir que le cyanure d’hydrogène est petit et apolaire.
L’endocytose est un exemple de transport en masse par lequel de grandes quantités de molécules sont transportées activement dans une cellule en utilisant une vésicule (endo signifiant dedans, cyto signifiant cellule). L’ ne se trouve probablement pas en quantités telles que l’endocytose soit nécessaire. Pour cette question particulière, on peut supposer que la concentration d’ dans les cellules de l’herbivore est beaucoup plus faible qu’à l’extérieur, car la molécule est toxique en inhibant l’un des processus de la respiration. Cela suggérerait un transport passif mais, puisque toutes nos options pourraient être passives en dehors de l’endocytose, cela ne nous aide pas à affiner nos options au-delà de l’élimination de l’endocytose.
Les molécules de grande taille ou polaires ne peuvent pas diffuser directement à travers les membranes et doivent donc utiliser un transporteur membranaire ou un canal ionique. Cette molécule est à la fois petite et apolaire, ce qui suggère qu’elle ne nécessite pas de protéine et qu’elle peut simplement diffuser à travers les phospholipides.
Notre réponse correcte est donc la A : elle diffuse à travers la bicouche lipidique.
La température a un impact significatif sur la fluidité des membranes plasmiques.
À basses températures, les phospholipides ont tendance à moins se déplacer et à s’organiser plus étroitement en une structure cristalline. Par conséquent, les basses températures rendent les membranes plus rigides, ce qui peut interférer avec des fonctions clés comme le passage des gaz. Une plus grande rigidité rend également les membranes plus cassantes. Le cholestérol limite ce phénomène en perturbant l’organisation serrée et régulière des phospholipides avec sa queue volumineuse, ce qui augmente la fluidité de la membrane.
À des températures plus élevées, plus d’énergie thermique (chaleur) est fournie aux phospholipides dans la bicouche, ce qui augmente leur énergie cinétique, ou vitesse de mouvement, et la fluidité de la membrane. Si la membrane est plus fluide, sa perméabilité augmente, car il y a plus d’espaces à travers lesquels les substances peuvent se déplacer. Un autre problème est que les protéines intégrées peuvent se dénaturer (changer de forme de manière irréversible) si la température dépasse leur niveau optimal. Cela signifie qu’elles ne peuvent plus remplir leurs fonctions de transport.
À des températures plus élevées, cependant, la queue de cholestérol maintient des interactions fortes avec les phospholipides et les resserre, ce qui empêche la membrane de devenir trop liquide. En conclusion, le cholestérol agit comme un tampon qui maintient une fluidité stable de la membrane sur un large éventail de températures.
Les solvants organiques moins polaires que l’eau, tels que les alcools ou les solvants apolaires, sont capables de dissoudre les membranes plasmiques. Prenons l’exemple d’un antiseptique : l’alcool perturbe les membranes des cellules bactériennes en dissolvant les acides gras des phospholipides, tuant ainsi la bactérie. Un alcool plus faible est présent dans les boissons alcoolisées, ce qui affecte les membranes cellulaires des neurones. Il perturbe la transmission de l’influx nerveux et ralentit les réactions de la personne, entre autres effets secondaires sur le comportement. Certaines substances plus hydrophobes, telles que certains médicaments, sont également capables de traverser directement la membrane.
Exemple 4: Décrire les conditions qui endommagent les membranes cellulaires
Quelle condition parmi les suivantes est la plus susceptible d’endommager une membrane cellulaire et d’augmenter sa perméabilité ?
- basses températures (4– )
- solutions hypertoniques
- solvants organiques
- solvants polaires
Réponse
Les températures élevées augmentent la perméabilité d’une membrane cellulaire en augmentant l’énergie cinétique et donc la fluidité des phospholipides dans la membrane, la rendant plus « fluide ». Les températures très élevées peuvent également augmenter la perméabilité en dénaturant de manière irréversible les protéines impliquées dans le transport de substances à travers la membrane, de sorte qu’elles ne puissent plus fonctionner correctement.
Les solvants organiques augmentent également la perméabilité d’une membrane cellulaire en dissolvant les phospholipides dans la membrane. C’est le cas pour tous les solvants moins polaires que l’eau, car ils ne peuvent pas interagir avec les phospholipides dans la bicouche de la même manière que les molécules d’eau polaires, donc ils ne peuvent pas maintenir la forme de la bicouche.
Les solutions hypertoniques (hyper, qui ont une forte concentration de soluté) ou hypotoniques (hypo, qui ont une faible concentration de soluté) peuvent avoir un impact sur la vitesse de déplacement des molécules de soluté et/ou d’eau à travers la membrane plasmique, mais cela ne sera pas susceptible d’affecter de manière drastique la perméabilité de la membrane.
Par conséquent, la condition qui influence le plus probablement l’augmentation de la perméabilité de la membrane cellulaire est la C : les solvants organiques.
Certaines expériences peuvent être menées pour étudier les effets de la température et de la concentration en solvant organique sur la structure et la perméabilité de la membrane. De telles expériences impliquent généralement un procédé appelé la colorimétrie, une technique utilisée pour mesurer la concentration de composés colorés ou de pigments dans une solution.
La colorimétrie est particulièrement utile lorsque les cellules expérimentales contiennent un pigment très concentré. Les cellules de betterave contiennent de la bétalaïne, un pigment qui leur donne une couleur rouge foncé distincte, et sont souvent utilisées en colorimétrie. La figure 6 ci-dessous illustre le fonctionnement de la colorimétrie.

Terme clé : Colorimètre
Un colorimètre compare la quantité de lumière traversant un échantillon de solution à la quantité de lumière qui peut traverser un échantillon de solvant pur et entièrement transparent.
Terme clé : Absorbance de la lumière
L’absorbance est une mesure de la quantité de lumière absorbée par un échantillon de solution dans un colorimètre.
Lorsqu’elles sont placées dans une solution aqueuse, une partie du pigments rouges s’échappe des cellules de betterave à travers leur membrane vers la solution. Plus la membrane cellulaire est perméable, plus le pigment fuira. La solution est ensuite versée dans un flacon carré appelé une cuvette, qui est placée dans un colorimètre (voir l’échantillon rouge sur la figure 6).
Le colorimètre est un appareil qui fait passer la lumière à travers une solution et mesure le pourcentage de lumière qui a été transmis et le pourcentage absorbé par le pigment. Si toute la lumière traverse un échantillon, c’est qu’aucune lumière n’a été absorbée par le pigment, donc l’absorbance sera nulle et la transmission au détecteur représentée figure 6 sera de .
Des filtres de couleur spécifiques sont généralement sélectionnés en fonction de la couleur de l’échantillon étudié, et placés entre la source de lumière et l’échantillon pour donner des valeurs d’absorbance par le pigment plus précises.
Le graphique de la figure 7 ci-dessous montre des exemples de résultats de cette expérience. Vous pouvez voir que lorsque la température augmente, l’absorbance de la lumière bleue par le pigment rouge bétalaïne augmente.

Récapitulons certains des points clés que nous avons abordés dans cette fiche explicative.
Points clés
- Les membranes plasmiques entourent chaque cellule vivante, et les eucaryotes ont aussi des membranes intracellulaires autour de certains de leurs organites pour la compartimentation.
- Le modèle de la mosaïque fluide décrit comment la membrane plasmique est constituée de phospholipides se déplaçant librement, à têtes polaires hydrophiles et queues d’acides gras hydrophobes, ainsi que de protéines et de cholestérol.
- La température affecte la perméabilité de la membrane en augmentant la fluidité des phospholipides et en dénaturant les protéines intégrées.
- Les solvants organiques, comme les alcools, affectent également la perméabilité de la membrane en dissolvant les phospholipides, ce qui la rend plus fluide et perméable.
- La perméabilité de la membrane peut être étudiée à l’aide d’un colorimètre pour mesurer l’absorbance de la lumière à travers une solution dans laquelle un pigment a traversé une membrane.

